Mikä on ontogeneesi biologian määritelmässä? Ongeneesin käsite
Ontogeneesi on organismin yksilöllinen kehitys alusta alkaen kuolemaan. Ontogeneesin ajanjaksoa hedelmöitetystä munasta nuoren yksilön vapautumiseen munakalvoista tai äidin kehosta kutsutaan alkion (alkion) kehitykseksi (alkion kehitys). Syntymisen tai munasta hautumisen jälkeen postembryoninen ajanjakso alkaa.
Eläinmaailmassa kolme yleisintä ontogeneesin tyyppiä ovat toukka, ei-toukka ja kohdunsisäinen. Ensimmäisessä niistä kehon kehitys tapahtuu muodonmuutoksen kanssa, toisessa muodostuminen tapahtuu munassa ja jälkimmäisessä äidin organismin sisällä.
Ihmisen alkiogeneesi on jaettu jaksoihin, joille jokaiselle on ominaista rakenteelliset piirteet, ravitsemustyyppi, hengitys ja erittyminen. Lääketieteellisessä käytännössä käytetään yleensä kahta käsitettä: Kahden viikon ajan kehittyvää organismia kutsutaan alkioksi (katso); alkaen 9. viikosta, toisin sanoen siitä, kun sikiö muodostaa elinten alkuvaiheet. Henkilön alkionjälkeinen jakso on jaettu ikäjaksoihin. Katso myös organogeneesi ,.
Ontogeneesi (kreikkalaisesta ontos - olemuksesta ja geneesistä - kehitys) on yksilöllisen kehityksen historia.
Charles Darwin antoi evoluutio-tulkinnan ontogeneesistä. Hän selitti eri lajien alkioiden samankaltaisuuden lajien välisten sukulaissuhteiden kanssa. Myöhemmin ontogeneesiä tutkittiin eri suuntiin. Alkionlehtien oppi säännölliseksi vaiheeksi kaikkien monisoluisten eläinten kehityksessä on luotu. Kokeellisen embryologian luominen antoi mahdolliseksi tutkia alkion kehitystä kokeellisesti. Viime vuosina fysiologinen ja biokemiallinen embryologia on kehittynyt menestyksekkäästi. Neuvostoliitossa embryologian ympäristösuunta alkoi kehittyä.
Monissa tapauksissa alkiotyypin tyypillä on tunnusomaista vuorottelua evoluution aikana: alkuperäisestä, primaarisesta vapaasta tourasta ei-toukkaan ja sitten yhdestä toissijaisen toukan alkionmuodosta.
Alkiogeneesin tyyppi liittyy sen periodisaatioon. Suurin jaksotusyksikkö on linkki. Useimmissa eläimissä ontogeneesi koostuu kolmesta linkistä. Jokainen linkki on jaettu useisiin jaksoihin. Joten, alempien selkärankaisten sukuside on jaettu kahteen jaksoon - alkio ja toukka, ylemmissä selkärankaisissa kolmeen - sukussa, esisikiöissä ja sikiössä. Myös siirtymäkauden tilat erotellaan - kuoriutuminen matelijoiden ja lintujen munankuorista; toimitus nisäkkäillä; monien eläinten metamorfoosi (katso).
Ihmisten ontogeneesin alkuvaiheille on ominaista nopeampi kehitysvauhti kuin sukulaisilla lajeilla (esimerkiksi simpanssit). Sitten tulee sekä synnytyksen että postnataalin kehityksen hidastuminen. Tämä hidastuminen johtuu aivojen voimakkaasta kasvusta sekä kohdunsisäisen kehityksen aikana (vastasyntyneen aivot painavat keskimäärin 340 g) että ensimmäisinä vuosina syntymän jälkeen (2-vuotiaan vauvan aivot painavat yli 1 kg). Aivojen kasvuun liittyy luutumisen hidastuminen osittain lantionrenkaan koosta riippuen.
Pienemmillä eläimillä on yleensä lyhyempi raskaus (joilla on läheinen suhde kahden istukkatyypin välillä).
Ihmisen postnataalinen ontogeneesi on jaettu neljään jaksoon (ikä): lapset, murrosikäiset, murrosikäiset ja aikuiset (kypsät).
Periodisaatio ja ontogeneesin tyypit.
I. Sikiökausi kehitys (kreikkalaisesta sanasta embryon - itä) -
Ensimmäiset 8 kehitysviikkoa: murskaava - blastulan yksikerroksisen alkion muodostuminen; gastrulaation - kahden ensimmäisen ja sitten kolmikerroksisen alkion muodostuminen - gastrula, tuloksena olevia kerroksia kutsutaan alkionlehdiksi; histogeneettisesti - kudoksen muodostuminen; organogeneesin - elinten muodostuminen.
Jokainen sukusoluista aiheuttaa yhden tai toisen elimen. alkaen ektodermi muodostunut: hermosto, ihon orvaskesi ja sen johdannaiset (kiimaiset vaa'at, höyhenet ja hiukset, hampaat). alkaen mesodermi lihakset, luuranko, erittyvä, lisääntymis- ja verenkiertoelimet muodostuvat. alkaen endodermi ruoansulatuskanava ja sen rauhaset (maksa, haima), hengityselimet muodostuvat.
I - tsygootti;
II - 2 blastomeeria;
II - 8 blastomeeria;
II - 32 blastomeeria (morula);
III - blastulan vaihe;
IV - gastrula;
V - kudosten ja elinten asettaminen:
1 - hermoputki;
2 - sointu;
3 - ektoderma;
4 - endodermi;
5 - mesodermi.
Kuva Laseletin kehityksen varhaiset vaiheet
Sikiön (sikiön) kehitysjakso. (sikiö - sikiö). Yhdeksännestä viikosta alkaen, kun alkiossa on jo kaikki elinjärjestelmät. Alkaen viikosta 9, ihmissikiö kutsutaan hedelmä . Ihmisillä synnytyksen kehitys kestää 38-42 viikkoa (kreikkalaisesta "ante" - ennen, "natus" - syntymästä)
II. postembryonic kehityskausi - syntymästä organismin kuolemaan.
Nuorten ajanjakso (puberteettiin saakka) etenee ontogeneesin tyypistä riippuen: suora tyyppi tai kehittyy metamorfoosin kanssa
suorakehitystyyppi - syntyneellä organismilla on kaikki aikuisen eläimen perusominaisuudet, se eroaa pääosin ruumiin koosta ja mittasuhteista. Korkeampien nisäkkäiden ja ihmisten kohdunsisäinen kehitys on tyypillistä, matelijoiden ja lintujen - muninta.
Poikkeus: munasarjoiset nisäkkäät - piikkikorko ja yachidna.
epäsuorakehitystyyppi - alkion kehitys johtaa toukan kehitykseen, joka eroaa aikuisen organismista ulkoisissa ja sisäisissä oireissa. Se on ominaista monille selkärangattomille, usein kaloille. Esimerkki: toukka kehittyy perhonen munista, ja juontokokot sammalamuneista.
 Toukan muuttumisen aikuismuotoksi ominaispiirteistä riippuen on olemassa 2 tyyppiä epäsuoraa ontogeneesiä:
Toukan muuttumisen aikuismuotoksi ominaispiirteistä riippuen on olemassa 2 tyyppiä epäsuoraa ontogeneesiä:
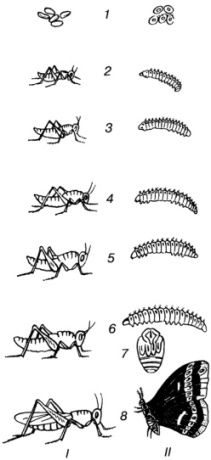
C epätäydellinen muutos - toukat kehittyvät vähitellen, menettäen peräkkäin väliaikaiset toukkaelimet ja saavuttaen aikuisen pysyvät ominaisuudet. Esimerkki: kurpitsat - elävät vesiympäristössä, niillä on väliaikaiset kidusselimet, häntä, kaksikammioinen sydän; aikuiset sammakot - keuhkot, 3-kammion sydän, raajat. Se on ominaista myös: punkkeille, lupille, ortopteraaneille (heinäsirkka, lice, sudenkorennot, torakat). kasvu- ja kehitysprosessissa toukat sulaavat useita kertoja (torakat sulaavat 6 kertaa) ja jokaisen pistoksen jälkeen muuttuvat yhä enemmän aikuiseksi.
C täydellinen muutos (metamorfoosi ) on ominaista useille hyönteisten, perhosten, bugien, dipteranien (hyttysiä, kärpäsiä), hymenoptera (mehiläiset, ampiaiset, muurahaiset), kirput jne. Toukat ovat matomaisessa rakenteessa ja ovat täysin erilaisia \u200b\u200bkuin aikuiset.
Kuva Hyönteisten kehitys, joiden lakkaaminen on epätäydellistä (I) ja täydellistä (II). 1 - munat, 2,3,4,5,6 - toukat; 7 - pupa; 8 - aikuisten muoto (imago).
Ruokintajakson lopussa toukka muuttuu liikkumattomaksi vaiheeksi - kotelo peitetty tiheällä kitiinikotelolla. Pupun sisällä erityiset entsyymit hajottavat kaikki elimet, lukuun ottamatta useita soluja, joita kutsutaan mielikiertoiksi. Aikuisten elimet kehittyvät kiekkosoluista.
Aikuinen murrosikä. Sille on ominaista suurin riippumattomuus, organismin aktiivisuus ympäristössä.
Vanhuuden ajanjakso.
Kasvu ja kehitys.
Toiminnallisten järjestelmien siirtymiselle kehon kypsymismuodolle on ominaista kehon elinten ja kudosten kasvu, sopivien kehosuhteiden asettaminen. Yksilöllisen kehityksen prosessissa erotetaan useita kasvutyyppejä: rajoitettu ja rajoittamaton; isometrinen ja allometrinen.
rajallinen (Määritetään). Kasvu rajoittuu tiettyihin ontogeneesin vaiheisiin. Esimerkki: hyönteiset kasvavat vain moltin aikana; ihmisillä kasvu pysähtyy 13-15-vuotiaana. Puberteettien aikana saattaa olla murrosiän kasvuleikkaus.
rajoittamaton kasvua havaitaan kaloissa, sisäkasveissa koko elämän ajan tai monivuotisissa kasveissa.
Isometrinen kasvu - kasvu, jolla elin kasvaa samalla vauhdilla kuin muu vartalo. Kehon koon muutos ei seuraa sen muodon muutosta. Kalalle ja hyönteiselle ominainen, jos se on epätäydellisesti muuttunut (heinäsirkat, siipiä ja sukuelimiä lukuun ottamatta)
allometrisiä kutsutaan kasvuksi, jolla annettu elin kasvaa tällä vauhdilla kuin muu vartalo. Kehon kasvu johtaa muutoksiin sen mittasuhteissa. Ominainen nisäkkäille ja ihmisille. Lähes kaikilla eläimillä on viime kädessä sukuelinten kehitys.
GENEETIKAN PERUSTEET.
genetiikka - Tiede, joka tutkii perintö- ja vaihtelulakeja.
Genetiikan tehtävä: perinnöllisen tiedon varastoinnin, siirtämisen, vaihtelevuuden toteuttamisen ongelmien tutkiminen.
menetelmät:
1. Hybridi-menetelmä (ristit) - kehittänyt G. Mendel, on tärkein geenitutkimuksessa. Menetelmä antaa mahdollisuuden paljastaa yksittäisten hahmojen perintökuviot ja ominaisuudet organismien seksuaalisen lisääntymisen aikana.
2. Sytogeneettinen menetelmä - antaa sinun tutkia kehosolujen kariotyyppiä ja tunnistaa genomi- ja kromosomimutaatiot. Tämän menetelmän myötä monien ihmisten sairauksien syyt on selvitetty (s. P. Down).
3. Genealoginen menetelmä (sukutaulut) - piirteiden perintötutkimukset ihmisellä useiden sukupolvien ajan (sukutaulu laaditaan, perheenjäsenet, joilla on tutkittava piirre, kirjataan)
4. Twin-menetelmä - tutkia kaksosia, joilla on samat genotyypit, sadan avulla voit tunnistaa ympäristön vaikutukset hahmojen muodostumiseen.
5. Biokemiallinen menetelmä - tutkii geenimutaatioista johtuvia aineenvaihduntahäiriöitä.
6. Väestötilastollinen menetelmä - avulla voit laskea geenien ja genotyyppien esiintymistiheyden populaatiossa.
Peruskäsitteet.

Ulostukseen liittyy läheisesti toisiinsa liittyvä risti.
Ontogeneesi on yksilön yksilöllisen kehityksen prosessi tsygootin muodostumisen hetkestä seksuaalisen lisääntymisen aikana (tai tytär-yksilön syntymiseen aseksuaalisen aikana) elämän loppuun.
Ontegeneesin periodisointi perustuu yksilön mahdollisuuteen sukupuolielimiin. Tämän periaatteen mukaan ontogeneesi on jaettu kolmeen jaksoon: lisääntymisen esto, lisääntyminen ja lisääntymisen jälkeinen jakso.
Esiintymistä edeltävälle ajanjaksolle on ominaista yksilön kyvyttömyys lisääntyä sukupuolensa vuoksi sen epäkypsyydestä. Tänä aikana tapahtuu anatomisia ja fysiologisia perusmuutoksia, jotka muodostavat seksuaalisesti kypsän organismin. Lisääntymistä edeltävällä ajanjaksolla henkilö on alttiimpi fysikaalisten, kemiallisten ja biologisten ympäristötekijöiden haitallisille vaikutuksille.
Tämä ajanjakso puolestaan \u200b\u200bjaetaan neljään jaksoon: alkion, toukka, metamorfoosi ja nuoruusiä.
Alkion (alkion) jakso kestää munan hedelmöityshetkestä alkion poistumiseen munakalvoista.
Toukkakausi esiintyy joillakin ala-selkärankaisten eläinten edustajilla, joiden alkioilla, jotka ovat poistuneet munakalvoista, on jonkin aikaa olemassa ilman kaikkia kypsän yksilön piirteitä. Touralle ovat ominaisia \u200b\u200byksilön alkion piirteet, väliaikaisten apuelinten läsnäolo ja kyky ruokkia ja lisääntyä aktiivisesti. Tämän ansiosta toukka viimeistelee kehityksensä suotuisimmissa olosuhteissa.
Metamorfoosille ontogeneesin ajanjaksona on ominaista yksilön rakenteelliset muutokset. Tässä tapauksessa apuelimet tuhoutuvat ja pysyvät elimet paranevat tai muodostetaan uudelleen.
Nuorten kausi kestää hetkestä, kun metamorfoosi loppuu, kunnes se tulee lisääntymisjaksoon. Tänä aikana yksilö kasvaa nopeasti, tapahtuu lopullinen elinten ja järjestelmien rakenteen ja toiminnan muodostuminen.
Lisääntymisjaksolla yksilö tajuaa kykynsä lisääntyä. Tänä kehitysjaksona se on lopulta muodostunut ja kestävä haitallisille ulkoisille tekijöille.
Lisääntymisen jälkeinen aika liittyy kehon asteittaiseen ikääntymiseen. Sille on ominaista lisääntymistoiminnan heikkeneminen ja sitten täydellinen häviäminen, käänteiset rakenteelliset ja toiminnalliset muutokset elimissä ja kehon järjestelmissä. Kestävyys erilaisille haittavaikutuksille on vähentynyt.
Postembryoninen kehitys voi olla suoraa ja epäsuoraa. Aikuisen kaltainen organismi, jolla on suora (ilman toukkia) kehitys, poistuu munakalvoista tai äidin kehosta. Näiden eläinten postembryoninen kehitys pelkistyy pääasiassa kasvuun ja murrosikään. Suoraa kehitystä tapahtuu eläimillä, jotka lisääntyvät munivilla munilla, kun munissa on runsaasti keltuaista (selkärangattomat, kalat, matelijat, linnut, jotkut nisäkkäät) ja eläinperäisissä muodoissa. Jälkimmäisessä tapauksessa munista puuttuu melkein keltuainen. Alkio kehittyy äidin kehossa ja sen elintärkeä aktiivisuus tapahtuu istukan (istukan nisäkkäiden ja ihmisten) välityksellä.
Epäsuora kehitys on toukkia, joissa on muodonmuutos. Metamorfoosi voi olla epätäydellinen, kun toukka muistuttaa aikuista organismia ja jokaisen uuden sulan ollessa yhä samankaltaisempi kuin se, ja täydellinen, kun toukka eroaa aikuisen organismista monien tärkeiden ulkoisen ja sisäisen rakenteen merkkien suhteen ja pupu vaihe on läsnä elinkaaressa.
esittely
Ontogeneesi on organismin yksilöllinen kehitys, jonka aikana sen morfofysiologiset, fysiologiset, biokemialliset ja sytogeneettiset ominaisuudet muuttuvat. Ontogeneesi sisältää kaksi prosessiryhmää: morfogeneesi ja lisääntyminen (lisääntyminen): morfogeneesin seurauksena muodostuu lisääntymiskypsä yksilö. Ontogeneesille on ominaista resistenssi - homeoressi. Kotitutkimus on stabiloitu tapahtumavirta, joka on prosessi organismin rakenteen, kehityksen ja toiminnan geneettisen ohjelman toteuttamiseksi.
Evoluution näkökulmasta tarkastellaan seuraavia ontogeneesin näkökohtia: alkion mukautumiset; filembriogenezy; ontogeneesin autonomisointi; ontogeneesin alkio.
Ongeneesin pääominaisuudet
Alkuohjelmoidut prosessit. Meioosin ja hedelmöityksen seurauksena muodostuneen ainutlaatuisen, muuttumattoman geneettisen kehitysohjelman läsnäolo
Ongeneesin peruuttamattomuus. Geneettistä ohjelmaa toteutettaessa on mahdotonta palata edellisiin vaiheisiin
Syventyvä erikoistuminen: Kehityksen myötä ontogeneesin polun muutoksen todennäköisyys vähenee
Adaptiivinen luonne: ontogeneesin monimuuttuja antaa kyvyn mukautua erilaisiin olosuhteisiin
Epätasainen tahti: kasvun ja kehityksen nopeus muuttuu.
Yksittäisten vaiheiden eheys ja jatkuvuus. Myöhemmissä vaiheissa ilmestyvät merkit perustuvat alkuvaiheessa näkyviin merkkeihin.
Syklisyyden läsnäolo: siellä on ikääntymisen ja nuorentamisen kierto
Kriittisten ajanjaksojen esiintyminen, jotka liittyvät polun valintaan solmupisteissä (harmaistuspisteissä) tai energian kynnysarvojen ylittämisessä.
Pääasialliset ontogeneesityypit
1. Organismien ontogeneesi, joilla on lisääntymiskyky ja / tai joilla on tsygoottinen meioosi (prokaryootit ja jotkut eukaryootit).
2. Sellaisten organismien, joilla on vuorottelevat ydinfaasit, itiögeneesi itiöiden meioosissa (useimmat kasvit ja sienet).
3. Organismien ontogeneesi vuorottelevalla seksuaalisella ja aseksuaalisella lisääntymisellä muuttamatta ydinfaaseja. Metageneesi on sukupolvien vaihtamista suolistossa. Heterogony on partenogeneettisten ja amfimiktisten sukupolvien vuorottelu matoissa, joissain niveljalkaisissa ja alemmissa chordateissa.
4. Ontogeneesi toukokuiden ja välivaiheiden kanssa: primaarisesta toukka-anamorfoosista täydelliseen metamorfoosiin. Koska munassa ei ole ravinteita, toukkavaiheet mahdollistavat morfogeneesin loppuun saattamisen ja joissain tapauksissa myös yksilöiden uudelleensijoittamisen.
5. Ontogeneesi ja yksittäisten vaiheiden menetys. Toukkavaiheiden ja / tai epäseksuaalisen lisääntymisen vaiheiden menetys: makean veden hydrat, oligochaetes, useimmat mahalaukun nilviäiset. Viimeisten vaiheiden ja lisääntymisen menetys ontogeneesin varhaisissa vaiheissa: neoteny.
Siksi on olemassa monia perustyyppejä ontogeneesistä ja vielä suurempi määrä johdettuja tyyppejä. Evoluutioteoria harkitsee ontogeneesiä yleensä kukinnan kasvien ja selkärankaisten avulla.
Kun otetaan huomioon ontogeneesin evoluution morfologinen puoli, joka on paleontologille välttämättömämpi, on ensinnäkin pidettävä mielessä suhde yksilöllisen kehityksen muutosten ja aikuisen organismin evoluution välillä. Juuri tämä ongelma muodostaa esimerkiksi A. N. Severtsovin fylembryogeneesin koko opin ytimen. Vaikka tällaista huomiota kehitystyön tuloksiin pidetään joskus jonkinlaisena Haeckelin ajattelun jäännöksenä tai parhaimmillaan hyväksyttävänä metodologisena välineenä, todellisuudessa se on väistämätöntä, jos olemme kiinnostuneita todellisesta evoluutioprosessista. Vain eristetyssä ontogeneettisessä jaksossa aikuisen vaihe voi vaikuttaa vähiten merkitsevältä, koska se on täysin määritetty aikaisemman kehityksen perusteella, mutta se ei itse määrittele mitään. Evoluutiossa asia esitetään aivan eri tavalla. Aikuisvaihe on ainoa, jossa normaali toiminta (ainakin Metazoa) on lisääntymistoiminto, joka luo geneettisen monimuotoisuuden, ts. Evoluutioprosessin materiaalin. Aikuisten organismien menestys taistelussa olemassaolosta riippuu siitä, millaisia \u200b\u200bsukusoluja käytetään seuraavan populaation sukupolven luomiseen, ja siksi siitä, millaiseen fenotyyppiseen materiaalivalintaan vastataan tulevaisuudessa. Valintaprosessissa epäonnistuneiden fenotyyppien ohella eliminoidaan niitä toteuttavat ontogeenit.
Tämä tarkoittaa, että riippumatta siitä, mitä onnistuneita mukautuksia luodaan kehitysvaiheen vaiheissa, ne eivät anna etuja, jos sen seurauksena ontogeneesi ei takaa halutun fenotyypin toteutumista. Aikuisvaihe, sellaisena kuin se oli, sanoo olosuhteet koko ontogeneesille. Siksi E. Haeckelin usein kritisoiduilla sanoilla, että fylogeenialla (jonka hän ymmärtää joukkona aikuisten vaiheita) on synnyn syy, on itse asiassa syvä merkitys, vaikkakaan ei aivan se, minkä biogeneettisen lain kirjoittaja väittää.
Se tosiasia, että kaikki ontogeneesin vaiheet muuttuvat, ei kukaan epäile. Kysymys on erilainen - miten niiden muutokset liittyvät? Kehittyykö aikuisaste itsenäisesti vai varhaisen kehityksen samanaikaisten morfologisten muutosten vaikutuksesta, vai molemmilla tavoilla kerralla? Viimeinkin, onko fylogeneesissä mahdollista kääntää myöhäisten muutosten vaikutukset edellisten vaiheiden kulkuun - toisin sanoen sellaista, jota ei voida ajatella yhden ontogeneesin puitteissa?
Ensimmäiseen kysymykseen annettu vastaus paljastaa ilmeisen ristiriidan olemassa olevissa näkemyksissä ontogeneesin roolista evoluutiossa, yhdistäen itse asiassa kaksi yhteensopimatonta käsitettä. Yhtäältä tunnustetaan, että normaali kehitys on säännelty prosessi, jolla pyritään tasaisesti lopulliseen tulokseen ja joka pyrkii mitätöimään kaikki tällä tiellä tapahtuvat poikkeamat. Tästä seuraa, että näiden variaatioiden ei pitäisi vaikuttaa suoraan aikuisen organismin ulkonäköön. Viimeksi mainitun kehitys tulisi liittää kehitysvaiheeseen liittyviin muodollisiin poikkeavuuksiin, joissa sääntelyn mahdollisuudet vähenevät. Ja samalla vallitsee usko, että muutokset alku- tai keskivaiheissa voivat aiheuttaa välittömän poikkeaman (poikkeaman) koko ontogeneesin kulusta, mikä muuttaa radikaalisti aikuisen muodon ulkonäköä jälkeläisissä. Yleensä tätä ristiriitaa ei edes huomaa. Molemmat käsitteet ovat olleet olemassa jo pitkään, ja ne ovat lähtökohtaisesti myös klassisessa morfologiassa, mutta vain ensimmäinen niistä. Kuten osoitetaan, se voi olla yhdenmukainen evoluutioteorian vaatimusten kanssa.
Perustan sille loivat K. M. Baerin kaksi empiiristä yleistystä. Yksi niistä koskee alkion vaihtelevuuden vähentämistä peräkkäisissä vaiheissa ( - k. M. Baerin lausunto korkeimmasta kehitystä sääntelevästä tavoitteiden asettamisen valvonnasta). Toinen on kuuluisa Bairen laki tai sukusolujen laki. Kuten C. Darwin on jo huomauttanut, tämä ilmiö tarkoittaa evoluutioteorian puitteissa, että selektiolla on taipumus muuttaa organismeja pääasiassa myöhemmässä iässä. Myöhemmin näitä ideoita tuki kehitysmekaniikka, joka osoitti kokeellisesti morfogeneettisten prosessien kyvyn itsesäätyä, minkä seurauksena aikuinen organismi on vakaampi kuin sen toteutusmenetelmä (ontogeneesissä tai uudistumisen aikana). Tämä yleistäminen, joka tunnetaan nimellä Ru-sääntö tai tasa-arvoisuuden periaate, vahvisti kehitysvaiheen varhaisen konservatiivisuuden osoittaen, että aikuisten organisaation tulee tapahtua ensisijaisesti muutosten myöhään tapahtuvassa kehitysvaiheessa.
Tällaisten ideoiden tärkein lähde oli kuitenkin E. Haeckelin tärkein biogeneettinen laki, joka on järjestely yksilöllisen kehityksen rinnakkaisuuden ja "olentojen tikkaiden" vanhan idean evoluutiokielen suhteen. Tämän lain (ts. Fylogeneesin lyhyen toistumisen ontogeneesissä) täytäntöönpanoa ei voida kuvitella muutoin kuin "kehityksen äärellisten muutosten perusteella.
Koska E. Haeckel oli kiinnostunut ontogeneesistä ei niinkään evoluutiossaan kuin fylogeneettisten rekonstruointimenetelmien hankkimisessa, emme löydä erityisiä selityksiä syylle tällaiseen muutosprosessiin, lukuun ottamatta lukuisia ja vähän sanottavia perinnöllisyyden ja sopeutumisen "lakeja". Siitä huolimatta biogeneettinen laki sisältää täysin integroidun teorian ontogeneesin kehityksestä, mikä viittaa kahteen kohtaan:
a) aikuisaste kehittyy lisäämällä uusia genees vaiheita ontogeneesiin, mikä varmistaa uudelleenkehityksen aikuisten esi-isien esiintymisen ontogeneesissä (palingenesis);
b) välivaiheilla on oma adaptiivinen evoluutio, mikä vääristää aikuisen vaiheen muunnoksien ontogeneettisen ennätön (koenogeneesi).
Biogeneettinen laki liittyy yleensä E. Haeckelin mekaanisesti myrkyllisiin näkemyksiin, toisinaan jopa väittämällä, että se kumottiin löytämällä Mendelian tekijät ja mutaatioprosessi. Todellisuudessa perinnöllisyyden kysymyksellä ei kuitenkaan ole ratkaisevaa merkitystä lain arvioinnissa. Itse asiassa riippumatta siitä, liittyykö evoluutioinnovaatioiden esiintyminen myöhäisissä somaattisissa muutoksissa, jotka välittyvät jollain tavalla sukusolujen kautta (Lamarckian käsite), tai näiden solujen välittömään muutokseen (perinnöllisten tekijöiden käsite), joka tapauksessa uusi kehityssykli täytyy väistämättä alkaa tsygoottimuunnoksilla, ts. olla erilainen vanhemmasta alusta alkaen.
E. Haeckel ymmärsi selkeästi tämän olosuhteen eikä pitänyt sitä biogeneettisen lain esteenä osoittaen, että tietyn kehityksen ajankohtana tapahtuneet ja vanhempien sukusolujen välittämät muutokset paljastuvat ensimmäistä kertaa saman vaiheen jälkeläisissä (”samanaikaisen perimisen laki”). ). Tässä hän seurasi suoraan C. Darwiniä, joka muotoili perinnöllisyyden periaatteen sopivassa iässä ja korosti, että perinnöllisen veropetosten esiintyminen sukusoluissa ja sen näkyvä esiintyminen jälkeläisten yksilöllisessä kehityksessä ovat kaksi eri asiaa. Siksi ei ole yllättävää, että determinanttien tai geenien tunnistaminen perinnöllisyyden kantajina ei estänyt monia tutkijoita tunnistamaan biogeneettisen lain tai ainakin evoluutiomuutosten yhteyden myöhäisiin vaiheisiin.
Päinvastainen näkemys ontogeneesin kehityksestä, joka mahdollistaa aikuisten organismien muutoksen välttämällä kehityksen varhaisvaiheissa, on peräisin E. Geoffrey Saint-Hilairesta ja F. Mullerista. Useimmiten niillä ja nykyaikaisilla kirjoittajilla on sama perustelu - normaalin ontogeneesin (tai homologisten elinten morfogeneesin) vertailu ja havaitun lopullisen ongeneettisen eron tunnistaminen (Baer-alkion divergenssi) todelliseen evoluutioprosessiin. Jos esimerkiksi homologisten elinten morfogeneesi ja B kahdessa eri muodossa osuvat vaiheeseen x, ja sitten eroavat, sitten väittävät, että ruumis B kotoisin poikkeamalla vaiheessa x . Me osoitamme tämän väitteen metodologisen epäjohdonmukaisuuden. Käsite tällaisen evoluutiopolun mahdollisuudesta tai väistämättömyydestä ei ole koskaan muodostanut kokonaisvaltaista teoriaa, toisin kuin Haeckelian teoria. Siitä huolimatta, 1900-luvun alkuun mennessä. heistä tuli hallitsevia.
Yksi syy tähän oli vertailevan embryologian tosiseikkojen kertyminen, jotka tutkijoiden mukaan eivät ole biogeneettisen lain mukaisia. Evoluution tunnistaminen varhaisilla kehityspoikkeamilla eliminoi teesin uudelleenkertoamisen ensisijaisesta väistämättömyydestä, kuitenkin kustannuksella, että heidän olemassaolostaan \u200b\u200btuli mysteeri. Toinen kohta Haeckelin näkemysten kritiikissä liittyi embryologien kasvavaan vakaumukseen siitä, että todellisen selityksen yksilöllisen kehityksen etenemiselle on etsittävä tutkittaessa sen välittömiä syy-tekijöitä eikä rakennettaessa spekulatiivisia historiallisia periaatteita, jotka korvaavat kokemuksen. Tämä lähestymistapa johti kokeellisen (syy-) embryologian syntyyn, joka paljasti sukusolujen rakenteen äärimmäisen monimuotoisuuden ja spesifisyyden erilaisissa organismeissa. Jälkimmäinen osoitti selvästi, että tsygootti on sama evoluution tuote kuin aikuisella organismilla ja että korkeamman organismin kehitysvaiheen alkuvaihe ei ole sen yksisoluisen esi-isän toistaminen. "Kananmuna ei vastaa enemmän fylogeneettisen ketjun alkuperäistä linkkiä kuin kana itse." Kaikki tämä johti vakuutukseen, että evoluutio tapahtuu muuttamalla koko ontogeneettistä sykliä ylhäältä alas jokaisessa seuraavassa sukupolvessa, eikä lisäämällä viimeisiä vaiheita. Ontogeny luo siten fylogenyyn eikä toista sitä. Samaan aikaan jotkut kirjoittajat pitivät mielessä vain asteittaisia \u200b\u200bmuutoksia, kun taas toiset - mahdollisuutta teräviin poikkeavuuksiin, jotka muuttavat äkillisesti aikuisen muodon.
Näiden näkemysten puitteissa kuitenkin kaikki teoreettiset mahdollisuudet esiintyä rinnakkaisten syntymistä ontogeneesin ja fylogeneesin välillä, huolimatta niiden olemassaolosta näennäisesti ilmeisiltä tosiasioilta, katosivat. He yrittivät kiertää tämän vaikeuden väittämällä, että sellaisilla rinnakkaisilla linjoilla on puhtaasti morfogeneettiset eikä historialliset syyt, ts. Että esi-ikäisen aikuisvaiheen voidaan pitää jälkeläisen ontogeneesissä vain sikäli kuin se muodostaa tarvittavan perustan seuraavien vaiheiden toteuttamiselle. Toinen selitys oli, että suurin osa näistä biogeneettisen lain käyttämistä esimerkeistä ei oikeastaan \u200b\u200bviittaa aikuisten alempien muotojen esiintymisen toistumiseen korkeammassa ontogeneesissä, vaan yksinkertaisesti yleisen kehitysvaiheen säilyttämiseen molemmissa, ts. Sukusolujen samankaltaisuuden ilmenemismuodoihin. , kuten K. M. Baer huomautti. Tätä väitettä, jota toistetaan jatkuvasti ja esimerkiksi myöhemmin, pidetään yleensä Haeckelin evoluutiomallin ratkaisevana kumotuksena. Mutta omituisen kyllä, he eivät huomaa, että ikäraudan uudelleenjärjestelyn aikana joka kerta alkuperäisestä vaiheesta, sukusolujen samankaltaisuuden säilymisestä tulee yhtä uskomatonta kuin uudelleenkokoonpanon ilmestyminen!
Kaiken kaikkiaan myöhempien tutkijoiden asema suhteessa biogeneettiseen lakiin näyttää kuitenkin olevan maltillisempi ja sallii mahdollisuuden erilaisiin synnyssä. Vastaavasti esitetään näiden polkujen tai moodien erilaisia \u200b\u200bluokituksia, joissa Haeckel-evoluutiomenetelmä myöhäisten laajennusten avulla vie tämän tai sen paikan - suhteellisen laajasta, esimerkiksi A. N. Severtsovin, vähäpätöiseen, jonka on kirjoittanut G. de Vir. Miksi ontogeneesi muuttuu eri tavoin eri tapauksissa, näitä näkemyksiä ei selitetä (lukuun ottamatta tavanomaista väitettä, jonka mukaan varhaiset muutokset ovat tapa luoda suuria systemaattisia ryhmiä). A. N. Severtsov huomauttaa suoraan, että vastaus tähän kysymykseen ei ole ollenkaan hänen tehtävänsä osa, joka on vain selvittää, miten evoluutio voi kulkea. Hän myöntää, että hänen mielenosoituksensa fylembryogeneesistä eivät riipu tietyn evoluutioteorian omaksumisesta.
Erottuvien moodien hyvin monimuotoisuus johtaa niiden kirjoittajia usein siihen johtopäätökseen, että kaikki loogisesti ajateltavat tavat muuttaa yksilöllistä kehitystä todella toteutuvat evoluutiossa. Tämä merkitsee sen tunnistamista, että ontogeneesin muutoksissa ei ole yleisiä malleja. Tällainen pettymys 100-vuotisen ongelman tutkimuksen tuloksena E. Haeckelin jälkeen ei yleensä ole yllättävää olosuhteissa, joissa se perustui pelkästään normaalin ongeneesin vertailuun, ottamatta huomioon evoluutiomateriaalin - ontogeneettisen vaihtelevuuden toteuttavien mekanismien analyysiä.
Toinen vahvistus tälle on S. Gouldin työ ontogeneesin ja fylogeneesin suhteesta, joka on laajin aiheesta aiheesta viime vuosikymmeninä. Menetelmät ovat tässä samat - evoluutiomuodot erotetaan ja arvioidaan, kuinka paljon ne heijastavat fylogeneesin kulkua. Yritykset yhdistää ne kahden tyyppisiin mukautuviin strategioihin (r ja 7 "C-valinta) eivät muuta ongelmaan lähestymistavan perinteistä luonnetta. Ontegeneesin kehitys jatkuu eristettyjen prosessien summana, joista jokainen etenee omissa erityisolosuhteissa. Kysymys niiden yhteisistä perussyistä on edelleen olemassa. syrjään.