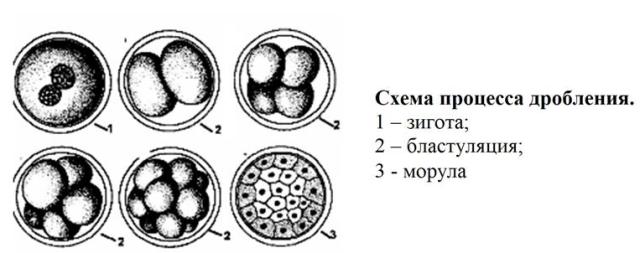
Ihmisen tsygootin jakautuminen. Zygote- ja murskausvaiheet
Antropologia ja biologian käsitteet Kurchanov Nikolay Anatolevich
Zygote- ja murskausvaiheet
Zygote- ja murskausvaiheet
tsygoottisukusolujen fuusiosta johtuva on monisoluisen organismin yksisoluinen kehitysvaihe. Vaikka tämän vaiheen kesto on yleensä pieni, on mahdollista jäljittää siinä tapahtuvat sytomorfologiset ja biokemialliset muutokset. Näillä muutoksilla on tärkeä rooli myöhemmissä alkiogeneesiprosesseissa. Monilla eläimillä proteiinisynteesi i-RNA: lla, joka muodostuu oogeneesin aikana, alkaa jo tsygootissa.
murskaavase on peräkkäisten mitoottisten jakautumisten prosessi, usein epätasainen. Murskauksen aikana muodostuneita soluja kutsutaan blastomeerejä. Koulutuksen murskaus päättyy blastulajolla on yleensä sisäinen onkalo - blastocoel. Murskausjakson ominaispiirre on kasvun puute. Vaikka blastula-vaiheessa oleva alkio voi koostua satoista soluista, sen koko ei ylitä tsygootin kokoa.
Munatyypistä riippuen, on olemassa erilaisia \u200b\u200bmurskaus- ja blastulatyyppejä. Munia on kahta päätyyppiä.
Homolekyyttiset munat - ytimen on oltava keskellä ja munankeltuainen on tasaisesti jakautunut sytoplasmassa.
Telolecital-munat - niillä on oltava selkeästi määritelty napaisuus, ytimen epäkeskoinen järjestely ja keltuainen, joka on jakautunut epätasaisesti sytoplasmassa.
Homolekyyttiset munat tuottavat yleensä räjähdyksiä, joissa on samat räjähdyssolut: tseloblastulu(ontelolla) tai morulaksi(ilman onteloa). Telolecital-munat antavat blastulon epätasaisilla blastomereillä: amfiblastulu(täydellinen murskaus) tai diskoblastulu(osittainen murskaus). Nisäkkäissä morula muodostuu sirpaloitumisen seurauksena, mutta silloin blastomeerien täydellisen asynkronisen jakautumisen aikana tapahtuu lisävaihe - sukusolu,tai blastokysti.
Kirjasta Psykofysiologian perusteet kirjoittaja Alexandrov Juri1.2. Muistin kiinnityksen vaiheet Hypoteesi kahdesta peräkkäin kehittyvästä jälkistä. Hypoteesin mukaan engrammin muodostaminen suoritetaan kahdessa vaiheessa: ensimmäiselle on tunnusomaista epävakaa jäljen muoto ja se on olemassa lyhyen ajan. Tämä on vaihe
Kirjasta Psykofysiologian perusteet kirjoittaja Alexandrov Juri3. HIDEN NUKEN JA NOPEEN NAPAN VAIHEET Lukuisten ja monipuolisten unetutkimusten vuosien aikana saamat tärkeimmät tiedot ovat seuraavat. Uni ei ole tauko aivojen toiminnassa, se on vain eri tila. Unen aikana aivot käyvät läpi useita eri vaiheita,
kirjoittaja Lerner George Isaakovich Kirjasta Biologia [Täydellinen opas tenttiin valmistautumiseen] kirjoittaja Lerner George Isaakovich Kirjasta Antropologia ja biologia käsitteet kirjoittaja Kurchanov Nikolay AnatolyevichFotosynteesin vaiheet Fotosynteesin prosessissa erotetaan kaksi vaihetta - vaalea ja tumma.Fotosynteesin valovaiheen aikana aurinkoenergiaa käytetään syntetisoimaan ATP: tä ja korkeaenergisia elektronikantoaineita. Minkä tahansa molekyylin absorboima valoenergia
Kirjasta Riittävän ravinnon ja trophologian teoria [taulukoita kuvilla] kirjoittaja Kirjasta Riittävän ravitsemuksen ja trofologian teoria [tekstitaulut] kirjoittaja Ugolev Alexander Mikhailovich9.2. Elämän alku ja kehitysvaiheet Nykyaikainen teoria elämän alkuperästä on teoria rakenteiden ilmestymisestä, jotka kehittyessä ja monimutkaistuessaan saavuttavat tietyt toiminnalliset ominaisuudet. Ilmeisesti nämä ominaisuudet olisivat voineet syntyä jo varhaisessa vaiheessa.
Materiaali otettu sivustolta www.hystology.ru
Murskaus on yksisoluisen tsygootin jatkokehitysprosessi, jonka aikana muodostuu monisoluinen blastula, joka koostuu seinämästä - blastodermistä ja onteosta - blastocoelista. Blastodermissa erotetaan katto, pohja ja niiden välinen reuna-alue. Tsygootin mitoottisen jakautumisen aikana muodostuu uusia soluja - blastomeereja, jotka pysyvät tiiviisti yhteydessä toisiinsa.
Murskauksen alkuvaiheessa monisoluinen organismi on kooltaan samanlainen kuin tsygootti, koska sen jakavat blastometrit eivät ole alkuperäisen solun kokoisia. Murskauksen luonne. evoluutiokorrasarja on erilainen, mikä johtuu suurelta osin munankeltuaisen keltuaisen määrästä ja jakautumisesta.
Murskaus voi olla täydellinen (holoblastinen) tai osittainen (meroblastinen). Holoblastisella murskaamisella kaikki tsygoottimateriaali osallistuu, meroblastisella murskaamisella, vain sen vyöhykkeellä, josta ei ole keltuaista.
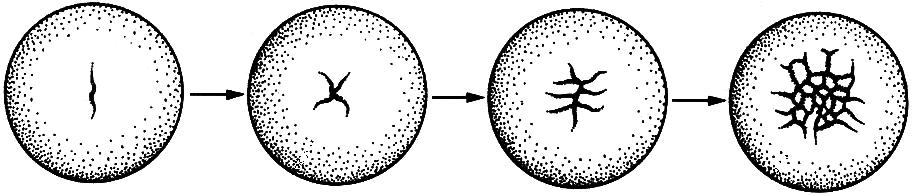
täysimurskaus luokitellaan tasaiseksi ja epätasaiseksi. Täydellinen yhtenäinen murskaus (kuva 43) on ominaista munille, joissa pieni määrä keltuaista (oligoletsiitaali) ja keltuaista (isoletsiitaali) jakautuvat tasaisesti koko solusytoplasmaan. Esimerkki tällaisesta murskaamisesta on lansetti, pyöreä mato jne. Hedelmöitetyssä munassa erotellaan kaksi napaa: ylempi - eläin ja alempi - vegetatiivinen.
Hedelmöityksen jälkeen keltuainen, jota pieni määrä oli tasaisesti jakautunut koko sytoplasmaan, siirtyy kasvillisuuteen. Murskauksen ensimmäinen ura kulkee meridiaalisuunnassa ja jakaa tsygootin kahteen räjäyttimeen, jotka vastaavat alkion rungon tulevaa vasenta ja oikeaa puolta. Murskauksen toinen ura kulkee myös meridionaalisesti suorassa kulmassa ensimmäiseen nähden, ja nyt alkio koostuu neljästä blastometristä. Kolmannella murskausuralla on päiväntasainen suunta, joten jokainen räjähdysmomentti on jaettu kahteen osaan. Tällainen alkio on rakennettu kahdeksasta blastomeeristä, kun taas neljä niistä muodostettiin tsygootin kasvullisesta navasta, ja siksi ne sisältävät koko sikotin keltuaisen ja ovat kooltaan suuria. Nämä räjähdysvaarat vastaavat vartalon takaosaa; eläin - neljä - etuosa.
Sitten ilmestyy kaksi meridionaalua, jakaen alkion 16 räjähdysryhmään. Viides murskaus on kaksi leveyttä uraa, alkiossa on 32 räjähdysmetriä. He alkavat
Kuva 43. Puristusuran järjestely lansetissa (A):
minä - alkio kahden blastomeerin vaiheessa; II - alkio neljän blastomeerin vaiheessa; III - alkio kahdeksan blastomeerin vaiheessa; IV - alkio 16 blastomeerin vaiheessa; V - alkio 32 blastomeerin vaiheessa; VI - alkio 64 blastomeerin vaiheessa; VII - alkio 128 blastomeerin vaiheessa. Blastularakenne (B): 1 - blastoderma; 2 - blastocele; 3 - pohja; 4 - reuna-alue; 5 - blastulan katto.
siirry vähitellen toisistaan \u200b\u200bkoskettamalla vain sivupintoja. Alkion sisään muodostuu ensin pieni onkalo - blastocele, joka vähitellen kasvaa. Kuudennen murskauksen jälkeen muodostuu 64 kennoa, kun murskausurat kulkevat meridionaalisesti. Seitsemännen sirpaloitumisen jälkeen (esiintyy neljä leveysuraa), alkio koostuu 128 blastomeeristä.
Myöhemmin alkion jakautumisen synkronointi katkeaa, blastomeerit siirtyvät reunaan ja sijaitsevat yhdessä kerroksessa muodostaen blastodermin, ja blastocele muodostuu alkion keskelle.
Murskaus päättyy blastulan muodostumiseen, jonka muoto muistuttaa nesteellä täytettyä palloa. Pallojen seinämän muodostavat blastodermisolut.
Siten täydellisen yhtenäisen murskaamisen yhteydessä koko tsygootin materiaali osallistuu jakoon ja jokaisen jaon (murskauksen) jälkeen solujen (blastomeerien) lukumäärä kaksinkertaistuu.
Blastodermissa erotellaan seuraavat osiot: katto, joka on rakennettu suhteellisen pienistä räjähdyssomeereista; pohja on suurempia blastomeerejä ja reunavyöhyke sijaitsee blastulan pohjan ja katon välillä.
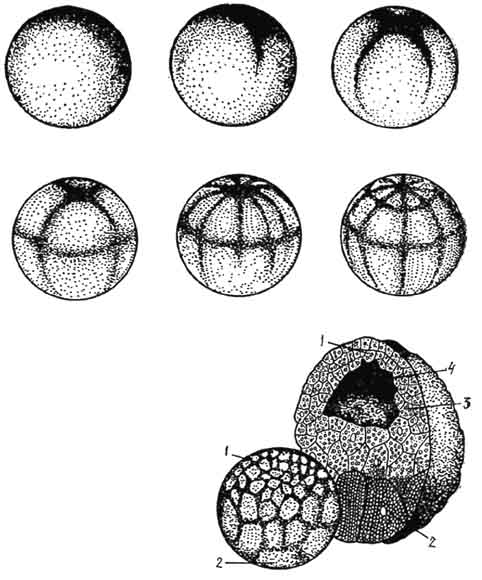
Kuva 44. Sammakkoeläinlääkkeen täydellinen epätasainen murskaus. Blastularakenne:
1 - mikrometriä; 2 - makrosyytit; 3 - blastoderma; 4 - blastocele.
Täydellinen epätasainen murskaus on ominaista mesolecitic (munankeltuaisen keskimääräinen määrä) ja telolecitic (munankeltuainen sijaitsee kasvillisuusnavassa) munille. Esimerkki tämän tyyppisestä murskaamisesta on sammakkoeläinsygoottien murskaus (kuva 44).
Murskaus alkaa muodostamalla kaksi puristusta, joka seuraa suorassa kulmassa meridionaalisia uria. Ne jakaa nopeasti tsygootin keltaiset eläinnavat kahteen ja sitten neljään pieneen blastomeeriin. Kasvillinen pylväs, joka sisältää koko tsygootin keltuaisen, murskataan paljon hitaammin, ja täällä syntyvät räjähdyssolut ovat suurempia.
Kolmas vako kulkee lähemmäksi tsygootin eläinnapaa ja siinä on leveyssuunta. Murskauksen leveysurat korvataan meridionaalisilla, ja asynkronisuus ja tangentiaalisuus (blastomeerien jakautuminen
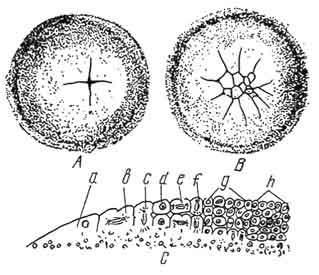
Kuva 45. Kanan itun osittainen (diskoidinen) pirstoutuminen:
A, B - murskausvaihe - ylhäältä päin (A - kaksi meridiona, - murskaamisen myöhemmässä vaiheessa) C - sukusolujen viillot (a, b, c, - keltuaiseen sijoittuneet marginaalisolut; d, e, f, g, h - keltuaista eristetyt solut).
taso tsygootin pinnan suuntaisena) sirpaloitumisessa, joten se päättyy monikerroksisen blastulan muodostumiseen. Blastulan katto on rakennettu pienistä blastomeereista, joita kutsutaan mikromereiksi. Pohja koostuu suurista blastomeereista - makromeereistä. Koko keltuainen on paikallistettu makromeereihin. Blastotsel siirtyi eläinnapaan ja pieneni kokoa. Holoblastisen (täydellisen) sirpaloitumisen aikana muodostunutta blastulaa kutsutaan koblastulaksi.
Osittainen tai meroblastinen (diskoidinen) murskausleviää kaloissa, matelijoissa, linnuissa ja on ominaista monisoluisille (monille keltuaisille) ja telolesiitaalisille munille (kuva 45).
Vain tsygootin eläinnavan pintakerros osallistuu murskaamiseen, koska solutuma ja sytoplasma ilman keltuaista sijaitsevat täällä. Loput tsygootista täytetään keltuaisella, joten sitä ei murskata.
Kaksi ensimmäistä meridionaa kulkevat eläinnavan läpi kulmassa toisiinsa. Ne eivät ulotu vegetatiiviseen napaan, ja siksi jälkimmäinen pysyy jakautumattomana blastomeereiksi. Meridionaalit antavat tien leveyteen ja tangentiaaliin. Murskauksen aikana muodostuneet räjähdyssolut sijaitsevat keltuaisessa yhdessä kerroksessa. Tätä kerrosta kutsutaan iturautalevyksi, siksi murskaamista kutsuttiin diskoidiseksi.
Alkion rungon rakentamiseen käytetään vain sen keskiosaa, sukusuojaa. Loput alkiolevystä osallistuvat väliaikaisten (väliaikaisten) elinten - ituskalvojen, muodostumiseen, jotka luovat suotuisat olosuhteet alkion kehittymiselle.
Hajanaisuus päättyy blastulan muodostumiseen, jossa blastosiila on kapean raon muodossa ja siirretään eläinnapaan. Blastulan katto on rakennettu blastomeereistä. Marginaalivyöhyke on ituradan levyn reunavyöhykkeen voimakkaasti jakavat solut (blastomeerit). Pohja on tsygoottisen vegetatiivisen navan keltuainen, jota ei ole jaettu blastomeereihin. Tämän tyyppistä blastulaa kutsutaan diskoblastulaksi.
Edellä olevasta materiaalista seuraa siten, että chordateilla on selvä yhteys munien keltuaisen määrän ja murskaamisen luonteen välillä. Se muuttuu täydestä (holoblastisesta) osaksi (meroblastiseksi) ja blastulasta - coeloblastulasta diskoblastulaan.
Kaikkien eläinluokkien kehittymisen alkioiden yhteiset ominaisuudet sirpaloitumisvaiheessa ovat solujen määrän ja siten myös DNA: n asteittainen lisääntyminen, koska tytärsolut ovat aina diploidisia; solupintojen pinta-alan kasvu; solupopulaatioiden alueellisten erojen lisääntyminen.
Sikotti (kreikkalainen sikottipari) on diploidi (sisältää täydellisen kaksinkertaisen joukon kromosomeja) solun, joka syntyy hedelmöityksestä (munan ja siittiön fuusio). Tsygootti on totipotentti (eli kykenee tuottamaan mitä tahansa muuta) solua. Termin esitteli saksalainen kasvitieteilijä E. Strasburger.
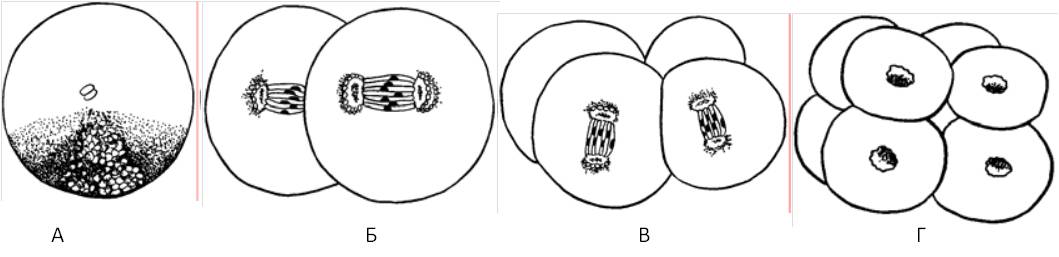
Ihmisillä tsygootin ensimmäinen mitoottinen jakautuminen tapahtuu noin 30 tuntia hedelmöityksen jälkeen, mikä johtuu monimutkaisista valmistusprosesseista ensimmäiseksi murskaamiseksi. Zygoottien murskaamalla muodostettuja soluja kutsutaan blastomeereiksi. Ensimmäisiä tsygoottijakautumisia kutsutaan murskaamiseksi, koska solu on täsmällisesti pirstoutunut: tytärisolut pienenevät kunkin jaon jälkeen, jakojen välillä ei ole solujen kasvuvaihetta.
Sikotin kehitys: Sikotti joko heti hedelmöityksen jälkeen alkaa kehittyä tai asettaa tiheän kuoren ja muuttuu jonkin aikaa lepääväksi itiöksi (jota usein kutsutaan zygosporeksi) - tyypillinen monille sienille ja levälle.
\u003e Murskaus
Monisoluisen eläimen alkionkehityskausi alkaa tsygootin pirstoutumisesta ja päättyy uuden yksilön syntymiseen. Murskausprosessi koostuu sarjasta peräkkäisiä tsygootin mitoottisia jakautumisia. Kahta solua, joka on muodostunut uuden tsygoottijakauman seurauksena, ja kaikkia seuraavia solusukupolvia tässä vaiheessa kutsutaan blastomeereiksi. Hajanaisuuden aikana yksi jakautuminen seuraa toista, eikä blastomeerejä muodostu, minkä seurauksena kutakin uutta blastomeerien sukupolvea edustavat pienemmät solut. Tämä solunjakautumisen ominaisuus hedelmöitetyn munan kehittymisen aikana määräsi myös kuviollisen termin - tsygootin murskaamisen - ilmestymisen.
Eri eläinlajeissa munat eroavat toissijaisesti ravintoravinteiden (keltuainen) jakautumisen määrästä ja luonteesta sytoplasmassa. Tämä määrää suurelta osin myöhemmän tsygootin murskaamisen luonteen. Pieni määrä ja keltuaisen jakautumista tasaisesti sytoplasmassa, koko tsygoottimassa jakautuu samojen blastomeerien muodostamiseen - täydellinen yhtenäinen murskaus (esimerkiksi nisäkkäillä). Kun keltuainen kerääntyy pääasiassa johonkin tsygoottipylväästä, tapahtuu epätasaista murskausta - muodostuu erikokoisia räjähdyksiä: suurempia makromeerejä ja mikromerejä (esimerkiksi sammakkoeläimiä). Jos muna on erittäin rikkaan keltuaisen kanssa, sen osa, keltaisesta vapaa, murskataan. Joten matelijoissa, linnuissa murskataan vain tsygootin kiekkomainen alue yhdessä navassa, jossa ydin sijaitsee, epätäydellinen, diskoidinen. Lopuksi, hyönteisissä, vain zygoottisen sytoplasman pintakerros osallistuu murskausprosessiin - epätäydellinen, pinnan murskaaminen.
Murskauksen tuloksena (kun halkeamien räjähtävien määrien lukumäärä saavuttaa merkittävän määrän) muodostuu blastula. Tyypillisessä tapauksessa (esimerkiksi lansetilla), blastula on ontto pallo, jonka seinämän muodostaa yksi solukerros (blastodermi). Blastulan onkalo on blastocele, jota muuten kutsutaan kehon primaariseksi onkaloksi ja joka on täytetty nesteellä. Sammakkoeläimillä blastulolla on hyvin pieni onkalo, ja joillakin eläimillä (esimerkiksi niveljalkaisilla) blastocele voi puuttua kokonaan.
Murskausvaiheen ydin. Murskaus - tämä on sarja peräkkäisiä tsygootin ja muiden blastomeerien mitoottisia jakautumisia, jotka päättyvät monisoluisen alkion muodostumiseen - blastula. Fragmentin ensimmäinen jakautuminen alkaa sen jälkeen kun ytimien perinnöllinen materiaali ja yhteinen metafaasilevy on muodostettu. Murskaamisesta saatuja soluja kutsutaan blastomeerejä (Kreikan kielestä blaste-itu, itu). Mitoosisten pilkkoutumisjakojen piirre on, että jokaisen jakautumisen myötä soluista tulee pienempiä ja pienempiä, kunnes ytimen tilavuuden suhde sytoplasmaan on normaali somaattisille soluille. Esimerkiksi merisiinissä tämä vaatii kuusi jakoa ja alkio koostuu 64 solusta. Peräkkäisten jakautumisten välillä solujen kasvua ei tapahdu, mutta DNA syntetisoidaan välttämättä.
Kaikki DNA-esiasteet ja tarvittavat entsyymit kerääntyvät munasolujen aikana. Tämän seurauksena mitoottiset jaksot lyhenevät ja jakautumiset seuraavat toisiaan paljon nopeammin kuin tavallisissa somaattisissa soluissa. Ensinnäkin, blastomeerit ovat vierekkäin, muodostaen soluklusterin, jota kutsutaan morulaksi. Sitten solujen väliin muodostuu ontelo - blastocoel, täynnä nestettä. Solut työnnetään reunaan, muodostaen blastulan seinämän - blastodermiin. Alkion kokonaiskoko fragmentoitumisen lopussa blastula-vaiheessa ei ylitä tsygootin kokoa.
Murskausjakson päätulos on tsygootin muutos monisoluinen yhden vaiheen alkio.
Murskauksen morfologia. Räjähdyssolut on yleensä järjestetty tiukassa järjestyksessä toistensa ja munan napa-akselin suhteen. Murskausjärjestys tai -menetelmä riippuu munan keltuaisen määrästä, tiheydestä ja jakautumisen luonteesta. Sachs-Gertwig -sääntöjen mukaan solun ytimellä on taipumus sijaita keltuaisen sytoplasman keskellä ja solun jakautumisen kara tämän vyöhykkeen suurimman suunnan suuntaan.
Oligo- ja mitacitic munissa, murskaaminen täydellinen, tai holoblastic. Tämän tyyppistä murskausta esiintyy nyrkkisummissa, joissain kaloissa, kaikissa sammakkoeläimissä, samoin kuin marsupiaaleissa ja istukkinisäkkäissä. Kun murskattu kokonaan, ensimmäisen jaon taso vastaa kahdenvälistä symmetriatasoa. Toisen jaon taso kulkee kohtisuorassa ensimmäisen tasoon nähden. Kahden ensimmäisen jaon molemmat vaunut ovat meridiaaneja, ts. Aloita eläinnavasta ja ulottuu kasvilliseen pylvääseen. Munasolu on jaettu neljään enemmän tai vähemmän samankokoiseen blastomeeriin. Kolmannen jaon taso ulottuu kohtisuorassa kahteen ensimmäiseen nähden leveyssuunnassa. Tämän jälkeen kahdeksan räjähdysaineen vaiheessa esiintyvissä mitescitic-munissa esiintyy epätasaista murskautumista. Eläinnavan kohdalla neljä pienempää blastomeeria - micromeres, kasvullisessa - neljä suurempaa - makromeerejä. Sitten jakautuminen tapahtuu jälleen meridiaanitasoilla ja sitten taas leveysasteella.
Luiden kalojen, matelijoiden, lintujen, samoin kuin yhden ohimennen nisäkkäiden monivalmisissa munasoluissa osittainen, tai merob-lasticheskoe, eli kattaa vain kellanmaton sytoplasma. Se sijaitsee ohuen kiekon muodossa eläinnavalla, joten tällaista murskaamista kutsutaan sidottuja.
Murskaustyyppiä karakterisoitaessa otetaan huomioon myös räjähdysrehujen suhteellinen sijainti ja halkeamisnopeus. Jos räjäyttimet ovat järjestetty riviin toistensa yläpuolelle sädettä pitkin, murskausta kutsutaan säteittäinen. Se on tyypillistä chordateille ja piikkinahkaisille. Luonnossa on muita muunnelmia blastomeerien alueellisesta järjestelystä sirpaloitumisen aikana, joka määrittelee sen tyypit, kuten spiraali nilviäisissä, kahdenväliset ascarisissa, anarkiset meduusissa.
Korrelaatiota havaittiin keltuaisen jakautumisen ja synkronoitumisasteen välillä eläin- ja kasvillisten räjähdysryhmien jakautumisessa. Oligolesiittisissa piikkinahkaisissa munissa pirstoutuminen on melkein synkronista, mesolecitic-munasoluissa synkronointi on heikentynyt kolmannen jaon jälkeen, koska vegetatiiviset räjähdyssolut, jotka johtuvat suuresta keltuaisten määrästä, jakautuvat hitaammin. Osittain pirstoutuneissa muodoissa jakautumiset ovat asynkronisia alusta alkaen ja keskiasennossa olevat blastomeerit jakautuvat nopeammin.
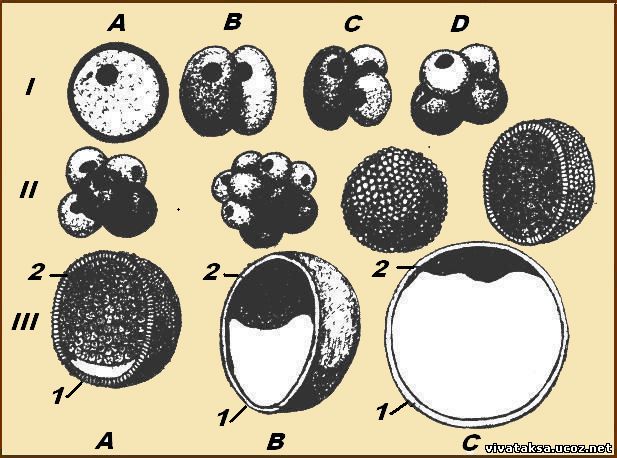
Kuva 7.2. Murskaus chordateissa erityyppisellä munalla.
A - lancelet; B - sammakko; B - siipikarja; G - nisäkäs:
minä- kaksi blastomeeria, II-neljä blastomeeria, IIIkahdeksan blastomeeria, IV-morulaksi, V-blastula;
1 murskausurat, 2 -blastomery, 3- blastodermiin, 4- blastoiel, 5- epiblast, 6- hypoblast, 7-embryoblast, 8- trofoblasti; kuvion ytimien koot eivät heijasta todellisia kokosuhteita
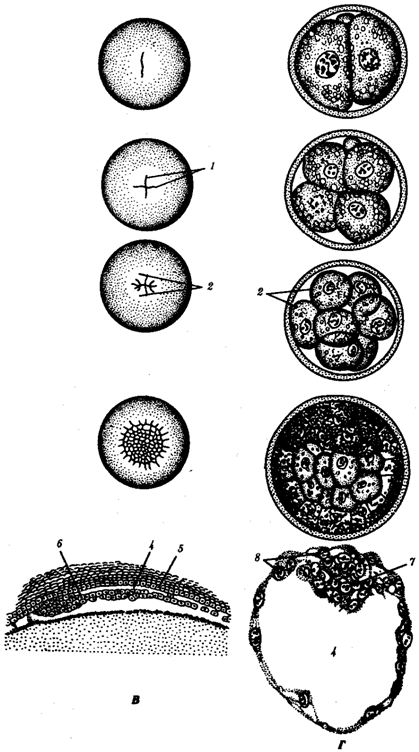
Kuva 7.2. laajentaminen
Murskauksen lopussa muodostuu blastula. Blastulan tyyppi riippuu sirpaloitumisen tyypistä ja siten munatyypistä. Joitakin murskaus- ja räjäytystyyppejä on esitetty kuvassa. 7.2 ja kaavio 7.1. Katso yksityiskohdat nisäkkäiden ja ihmisten murskaamisesta ks. 7.6.1.
Molekyylisten geneettisten ja biokemiallisten prosessien ominaisuudet murskaamisen aikana. Kuten yllä todettiin, mitoottiset syklit fragmentoitumisjaksossa lyhenevät huomattavasti, etenkin aivan alussa.
Esimerkiksi koko jakosykli merisiilin munissa kestää 30–40 minuuttia ja S-vaiheen kesto on vain 15 minuuttia. Gi- ja 02-jaksoja ei käytännössä ole, koska tarvittava määrä kaikkia aineita syntyy munasolun sytoplasmaan, ja mitä suurempi se on, sitä suurempi se on. Ennen kutakin jakautumista syntetisoidaan DNA ja histonit.
Replikaatiohaarukan etenemisnopeus DNA: n läpi fragmentin aikana on normaali. Samanaikaisesti blastomeeri-DNA: ssa on enemmän aloituskohtia kuin somaattisissa soluissa. DNA-synteesi tapahtuu kaikissa replikoneissa samanaikaisesti, synkronisesti. Siksi DNA: n replikaation aika ytimessä on yhdensuuntainen yhden, lisäksi lyhennetyn replikonin kaksinkertaistumisajan kanssa. On osoitettu, että kun ydin poistetaan tsygootista, tapahtuu sirpaloituminen ja alkio saavuttaa kehitysvaiheessaan lähes blastula-vaiheen. Jatkokehitys lakkaa.
Murskauksen alussa muun tyyppisiä ydinaktiivisuuksia, esimerkiksi transkriptio, puuttuu käytännössä. Erityyppisissä munissa geenitranskriptio ja RNA-synteesi alkavat eri vaiheissa. Tapauksissa, joissa sytoplasmassa on monia erilaisia \u200b\u200baineita, kuten esimerkiksi sammakkoeläimissä, transkriptio ei aktivoidu heti. Niissä RNA-synteesi alkaa varhaisessa blastula-vaiheessa. Sitä vastoin nisäkkäissä RNA-synteesi alkaa jo kahden blastomeerin vaiheessa.
Murskausjaksolla muodostuu RNA: ta ja proteiineja, samanlaisia \u200b\u200bkuin ne, jotka syntetisoidaan munasolujen aikana. Nämä ovat pääasiassa histoneja, solukalvojen proteiineja ja entsyymejä, jotka ovat välttämättömiä solun jakautumiselle. Edellä mainitut proteiinit käytetään välittömästi niiden proteiinien kanssa, jotka on aiemmin varastoitu munasolujen sytoplasmaan. Samanaikaisesti murskausjakson aikana on mahdollista proteiinien synteesi, jotka eivät aiemmin olleet mahdollisia. Tätä tukevat tiedot alueellisten erojen esiintymisestä RNA: n ja proteiinien synteesissä blastomeerien välillä. Joskus nämä RNA: t ja proteiinit alkavat toimia myöhemmissä vaiheissa.
Tärkeä rooli pirstoutumisessa on sytoplasman jakautumisella - sytokineesiin. Sillä on erityinen morfogeneettinen merkitys, koska se määrittelee pirstoutumisen tyypin. Sytotomian prosessissa supistus muodostetaan ensin käyttämällä mikrokuitujen supistuvaa rengasta. Tämän renkaan kokoonpano tapahtuu mitoottisen karan napojen välittömässä vaikutuksessa. Sytotomian jälkeen oligoleciitaalisten munien blastomeerit pysyvät kytkettynä vain ohuilla siltoilla. Juuri tällä hetkellä niitä oli helpoin erottaa. Tämä johtuu siitä, että sytotomia johtaa solujen välisen kosketuspinta-alan pienenemiseen kalvojen rajoitetun pinta-alan vuoksi.
Välittömästi sytotomian jälkeen solun pinnan uusien osien synteesi alkaa, kosketuspinta-ala kasvaa ja blastomeerit alkavat olla läheisessä kosketuksessa. Murskausurat kulkevat ovoplasman yksittäisten osien välisiä rajoja heijastaen ovoplasmisen erottelun ilmiötä. Siksi eri blastomeerien sytoplasman kemiallinen koostumus vaihtelee.
esittely
Lannoituksella on erittäin tärkeä rooli kehityksessä, mutta se on vasta sen ensimmäinen vaihe. Tsygootti, jolla on uusi geneettinen potentiaali ja uusi sytoplasman jakauma, perustaa monisoluisen organismin. Kaikissa tunnetuissa eläimissä tämä alkaa prosessilla, jota kutsutaan murskaava - Sarja mitoottisia jakautumisia, joiden seurauksena valtava tilavuus munasytoplasmasta jaetaan lukuisiin pienempiin soluihin, jotka sisältävät ytimen. Tällaisia \u200b\u200bmurskausjakson aikana muodostuneita soluja kutsutaan blastomeerejä.
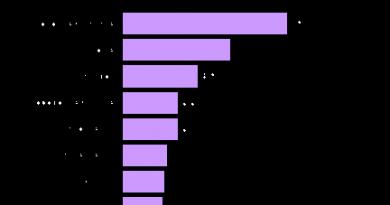
Useimmissa lajeissa alkion tilavuus ei kasva murskauksen aikana. Tämä on yllättävää, koska useimmissa muissa tapauksissa solujen lisääntymistä mitoosien välisellä ajanjaksolla tapahtuu solukasvu: solun määrä kaksinkertaistuu melkein ja jakautuu sitten. Tällainen kasvu johtaa selvästi lisääntyneeseen kokonaissolutilavuuteen pitäen samalla yllä ytimen tilavuuden suhde vakio-suhde sytoplasman tilavuuteen. Zygoottisen sirpaloitumisen aikana sytoplasman tilavuus ei kuitenkaan lisää: tsygootin sytoplasman valtava massa on jaettu yhä pienempiin soluihin. Ensin muna jaetaan kahteen puolikkaaseen, sitten neljään osaan, kahdekspaan osaan jne. Tämä munasytoplasman jakautuminen, johon ei liity kasvua, suoritetaan putoamalla jakautumisten välisestä kasvuvaiheesta, kun taas ydinjakaumat seuraavat toisiaan niin suurella nopeudella, jota ei koskaan havaita tulevaisuudessa (jopa kasvainsoluissa). Joten esimerkiksi sammakon muna voi jakaa 37 000 soluun 43 tunnissa, ja Drosophilassa murskauksen aikana mitooseja esiintyy joka toinen minuutti 2 tunnissa.
Murskauksen aikana solujen lukumäärän kasvuvauhti voidaan arvioida vertaamalla sitä niiden lukumäärän kasvun nopeuteen muissa kehitysvaiheissa. Kuvassa 1 Kuvio 3.1 esittää sammakkoalkion solumäärän logaritmin muutoksia ajan suhteen (Sze, 1953). Se kuvaa solunjakautumisen voimakkuuden voimakasta muutosta muutoksen aikana murskaamisesta ruoansulatukseen. Yksi murtumisen aikana tapahtuvan voimakkaan fissiovoiman seurauksista on sytoplasman tilavuuden ja ytimen tilavuuden suhteen asteittainen lasku. Muutos nopeudessa, jolla sytoplasman tilavuuden suhde ytimen tilavuuteen pienenee, monentyyppisissä ytimissä on ratkaiseva tekijä, joka määrittää joidenkin geenien aktivaatioajan.
sisään Xenopus laevis uusien matriisien transkription aktivaatio tapahtuu vasta 12 pilkkomisjaon jälkeen. Tässä vaiheessa pirstoutumisjakauman intensiteetti vähenee, blastomeerit saavat liikkuvuuden ja geenit alkavat transkriptoida ytimessä. Uskotaan, että blastomeerien pirstoutumisen voimakkuus riippuu jostakin tekijästä, jonka synteesi riippuu vasta muodostuneesta kromatiinista, koska siirtymäaikaa seuraavaan kehitysvaiheeseen voidaan siirtää muuttamalla ploidiaa : mitä enemmän kromatiinia alkio sisältää, sitä aikaisemmin siirtyminen tapahtuu. Jos kromatiinin määrä on kaksinkertainen normaaliin verrattuna, siirtymä on yhden jaon aikaisempi kuin normaali. Siten pirstoutuminen alkaa pian hedelmöityksen jälkeen ja päättyy, kun alkion sisällä saavutetaan uusi ydin ja sytoplasma.
Murskauksen käsite
Murskaus on sarja peräkkäisiä mitoottisia jakautumisia hedelmöitetystä tai kehityksessä aloitetusta munasta. Murskaus on alkion ensimmäisen kehityksen ajanjakso, joka esiintyy kaikkien monisoluisten eläinten ontogeneesissä. Tässä tapauksessa alkion massa ja sen tilavuus eivät muutu, pysyen samana kuin murskauksen alussa. Muna on jaettu yhä pienempiin soluihin - blastomeereihin. Murskauksen ominaispiirre on sytoplasman johtava sääntelyrooli kehityksessä. Murskauksen luonne riippuu keltuaisen määrästä ja sen sijainnista munassa. (Belousov L.V. 2005)
Murskauksen biologinen merkitys
siirtyminen monisoluisuuteen
lisääntynyt ytimen sytoplasminen suhde
Murskausjakso
päättyy kehon kehitysvaiheeseen, jota kutsutaanblastula Tsm.Tl7 Yksi murskausjakson tärkeimmistä piirteistä on p-tyypin kasvun puuttuminen. huolimatta pörssistäD shakshsh & sy daden ja minä Lisäksi murskaamisen päätyttyä alkion tilavuus vähenee jonkin verran. Aikuisten organismien solunjako liittyy yleensä läheisesti solujen kasvuun. Blastomeerien ontogeneesissä ei ole kasvujaksoa; blastomeerien koko ei kasva ennen pirstoutumista.Eläinten ytimen fragmentoituminen pienenee kooltaan, kun taas kunkin seuraavan fragmentoitumisen seurauksena oleva protoplasman tilavuus puolittuu. Siksi munan kuudennella tai seitsemännellä sirpaloitumisella (ja joillakin eläimillä neljännellä tai viidennellä), normaali tilavuussuhde palautetaan. Ytimet ja sytoplasmat blastomeereissä, jotka, kuten olemme nähneet, osoittautuvat vakavasti hajottamaan sukusoluja.
Tietyillä poikkeamilla odotetusta teoreettisesta diffraktiosta tällainen muutos ydin-plasma-suhteessa tapahtuu, kun kaikki munat murskataan. Esimerkiksi ennen merisiilimunaa hedelmöittämistä, ytimen tilavuuden suhde plasman tilavuuteen on 1: 550, ja murskauksen lopussa, ts. Blastula-vaiheessa, tämä suhde
1 : 6 . (Tokin B.P. 1987)MURASTUSMEKANISMI
Somaattisten solujen solusykli on jaettu neljään vaiheeseen (vaiheeseen). Mitoosia (M) seuraa presynteettinen ajanjakso (G1), jonka jälkeen tapahtuu DNA-synteesi (S). Sitten tulee premitoottinen vaihe (G2), jota seuraa taas mitoosi.
Blastomeerisolusykli fragmentoinnin varhaisvaiheissa voi olla paljon yksinkertaisempi - kaksivaiheinen. Tätä kaksifaasista sykliä ohjaavat tekijät paikallistuvat sytoplasmassa. Nämä ovat samoja tekijöitä, jotka säätelivät kypsymisjakaumia oogeneesin aikana: kypsyttämistä stimuloiva tekijä, sytostaattinen tekijä ja kalsiumionit.
On osoitettu, että jakautuvissa soluissa MPF-aktiivisuuden taso käy läpi syklisiä muutoksia. MPF-aktiivisuus sammakkoblastomeereissä fragmentin varhaisvaiheissa on korkein M-vaiheessa, eikä sitä havaita S-vaiheessa.
MPF: n toiminta suoritetaan muuttamalla ydinvaipan rakennetta. Sytostaattinen tekijä stabiloi kypsytymistekijää, pitäen solut mitoositilassa, ja kalsium inaktivoi sytostaattisen tekijän stimuloimalla S-vaiheeseen siirtymistä MPF: n inaktivoitumisen takia. Kun CSF: tä lisätään, myös sytoplasman kuoren kerroksen sykliset supistukset lakkaavat, ja myöhempi kalsiumionien injektio stimuloi niitä (kuvio 1).
Varhaisessa kehitysvaiheessa sytoplasma määrittelee solunjakautumisen nopeuden sekä S- ja M-vaiheiden keston. Asynkronisten murskausjaksojen aikana ilmestyy G1-vaihe, jakson kaikkien muiden vaiheiden kestoa pidennetään. Erityyppisten RNA: n synteesi DNA-matriiseissa alkaa, ts. alkion genomin transkriptioaktiivisuus herää.
Kuva 1. Blastomeerien solusyklin säätelymalli (Gilbert, 1993)
Geenit, jotka on tuotu alkion perimään spermasolun avulla, ilmentävät toimintaansa täsmällisesti tällä ajanjaksolla ja joka tapauksessa viimeistään synkronisen fragmentoitumisen ajanjakson lopussa. Tuolloin alkio otti kohtalon omiin käsiinsä ja lakkasi olemasta äidin geneettinen kopio. Koska asynkroninen ajanjakso alkaa eri määrän pilkkoutumisen jakautumisten jälkeen, transkriptioaktiivisuus herätetään vastaavasti erilaisella blastomeerien lukumäärällä: nisäkkäissä ja pyöreässä maassa melkein kehitysvaiheen alusta lähtien, piikkinahkoissa 32-blastomeerivaiheessa, sammakkoeläimissä blastula-vaiheesta.
Murskauksen ja munien luokittelu.
Munien luokittelu
Munat luokitellaan seuraavasti:
1. Keltuaisten lukumäärän perusteella - monisyytti, mesosyytti, oligosyytti ja alecital (ts. Monivyöhyke, puolikollainen, matala keltuainen ja ei-keltuainen).
2. Keltuaisen jakautumisen mukaan munan määrän mukaan:
Teloletsiitaali - keltuaisen määrä kasvaa epänormaalista navasta vegetatiiviseen;
Homolecital (isolecital) - vitelliinirakeet jakautuvat tasaisesti munapaksuuteen;
Centro lecital - vitelliinirakeista vapaa sytoplasma sijaitsee heti munankuoren alla, ytimen ympärillä, jolla on keskeinen sijainti, ja ohuina naruna, jotka yhdistävät nämä alueet. Välitila täytetään keltuaisella. (Knorre A. G)
Holoblastinen tyyppi - munien ja blastomeerien täydellinen erottaminen murskaamalla urat (a-, oligo-, mesolecitic, iso-, telolecital).
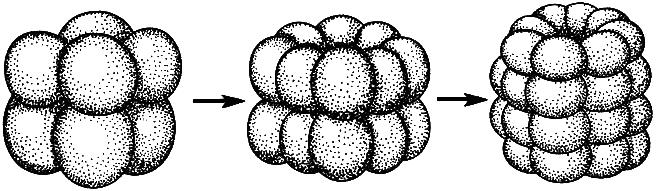
Piikkisolujen holoblastinen murskaus (Gilbert, 1993):
A, B, C - prosessin peräkkäiset vaiheet
2. Meroblastinen tyyppi - munan osittainen jakautuminen. Murskausurat tunkeutuvat syvälle munaan, mutta eivät erota sitä kokonaan. Keltuainen on jakautumaton:
Pintamurskaus (polysyyttiset, keskittymäiset munasolut) - sytoplasman pintakerroksen erottaminen yksittäisillä (aiemmin useaan kertaan jakautuneilla) ytimillä väliseinien avulla, jotka on suunnattu normaalisti munan pintaan. Munan keskiosa on jakautumaton:
Diskoidinen murskaus (polysykitaaliset, telelolekytaaliset munasolut) - vaurioita muodostuu ytimien fission jälkeen, mutta ne eivät jaa koko munaa, vaan vain yhden sen navoista.
Kananmunien diskoidinen murskaus (Gilbert, 1993):
Näkymä eläinnavan sivulta on esitetty. Murskausurat eivät ulotu keltuaiseen, ja tuloksena oleva blastoderma koostuu yhdestä solukerroksesta
3. Murskaamisesta saatujen määrien perusteella:
Tasainen - blastomeerien määrät ovat samat:
Epätasainen - blastomeerien määrät eivät ole samat.
4. Perustuen kario- ja sytotomian kestoon murskaaman munan eri blastomeereissä:
Synkroninen - murskaus alkaa ja loppuu kaikissa räjähdysaineissa samanaikaisesti;
Asynkroninen - jakautumisen alku ja aika eri blastomeereissä eivät ole samat.
5. Blastomeerien suhteellisen sijainnin perusteella murskaamassa:
Radiaalinen sirpaloituminen - blastomeerien suhteellinen sijainti on sellainen, että munan alkuperäinen napa-akseli toimii murskautuvan ytimen radiaalisen symmetrian akselina;
Spiraalimurskaus - murskaaman munan symmetrian asteittainen rikkominen, joka johtuu räjäytystesterien lopullisen jakautumisen spiraalisiirrosta toisiinsa nähden;
Kahdenväliset murskaukset - blastomeerit sijaitsevat siten, että alkion läpi voidaan vetää vain yksi symmetriataso;
Anarhistinen pirstoutuminen - kuvioiden puuttuminen blastomeerien sijainnista.
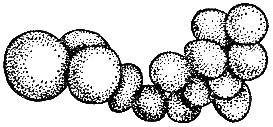
Anarhistinen pirstoutuminen (Tokinin mukaan, 1987)
Hajanaisuus päättyy blastulan muodostumiseen, monisoluiseen rakenteeseen, jonka sisällä on enemmän tai vähemmän ilmeinen onkalo. Jälkimmäistä kutsutaan murskausonkaloksi tai blastoceleksi. (Carlson B. M)
Murskaus.
Rannekkeen murskaus.
Pieni määrä keltuaista selittää murskaamisen ja mahalaukun helpon. Murskaus on radiaalityyppistä, melkein tasaista, ja seurauksena muodostuu koblastula.
Laselettimunan murskaus (Almazov, Sutulov, 1978):
A - tsygootti; B, C, D - blastomeerien muodostuminen
(näyttää jakokaran sijainnin)
Eläinnapa vastaa suunnilleen toukan rungon tulevaa etupäätä. Hedelmöittynyt muna (tsygootti) pilkotaan täysin blastomeereiksi oikeassa geometrisessa etenemisessä. Räjähdyssolut ovat melkein samankokoisia, eläimet ovat vain hiukan pienemmät kuin kasvulliset. Murskauksen ensimmäinen ura - meridinaali, kulkee eläin- ja kasvillisen navan läpi. Hän jakaa pallomaisen munan kahteen täysin symmetriseen puolikkaaseen, mutta räjähdysvaarat ovat pyöristettyjä. Ne ovat pallomaisia, niillä on pieni kosketusalue. Murskauksen toinen ura on myös meridionaalinen, per-
heilurin suhteessa ensimmäiseen ja kolmanteen - leveysleveyteen.
Kun blastomeerien lukumäärä kasvaa, ne eroavat yhä enemmän alkion keskustasta muodostaen keskelle suuren onkalon. Lopulta alkio on tyypillisen koblastulon muodossa - vesikkeli, jonka seinämä on muodostettu yhdestä solukerroksesta - blastodermi ja
ontelo, joka on täytetty blastocele-nesteellä.
Blastosolut, ensin pyöristetyt ja siksi eivät ole tiiviisti suljettuja, saavat prismien muodon ja sulkeutuvat tiukasti. Siksi myöhäistä blastuaalia, toisin kuin aikaista, kutsutaan epiteeliksi. Myöhäinen blastula-vaihe suorittaa murskausjakson. Tämän ajanjakson loppuun mennessä solukoko saavuttaa minimin, ja alkion kokonaismassa ei nouse verrattuna hedelmöitetyn munan painoon.
Murskaaminen sammakkoeläimissä.
Sammakkoeläinten pirstoutuminen on holoblastista (täydellistä), epätasaista ja asynkronista. Murskauksen ensimmäinen vako on meridionaalinen, kulkee eläinnavalta vegetatiiviseen. Koska sammakkoeläimen kasvillisuusnapa on ylikuormitettu keltuaisella, sitä lähempänä sitä, sitä nopeammin valan viilto pienenee. Hänellä ei vieläkään ole aikaa päästä vegetatiiviseen napaan, kun toinen vako alkaa kaatua.
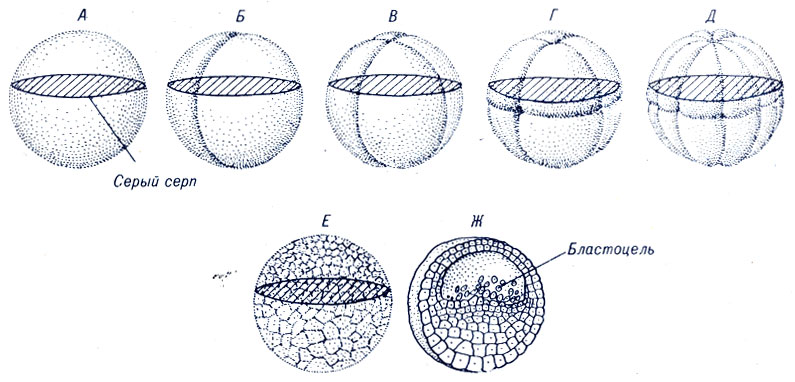
Kuva 3. Sammion (A - E) murskaaminen ja sammakon blastulan (G) poikkileikkaus
(Gilbertin, 1993 mukaan)
Toinen vako on myös meridionaalinen, kulkee kohtisuorassa ensimmäiseen nähden. Ensimmäisen varan ohittamisen jälkeen muodostuu kaksi räjähdyssomeeria, toisen jälkeen - neljä räjähdysmomeeria. Neljän blastomeerin vaiheessa kaksi eläimen blastomeeria saa ½ harmaata sirppimateriaalia, ja kahdessa vegetatiivisessa räjähdysaineessa ei saada harmaata sirppimateriaalia.
Kolmas murskausura on leveysleikkaus, joka kulkee lähemmäksi eläinnapaa. Sen läpikäynnin seurauksena muodostuu 8 räjähdysainetta: 4 eläin- ja 4 kasvillista, jotka eroavat toisistaan \u200b\u200bmerkittävästi. Seuraavat kaksi meridionaalua. Blastomeerien koon ja keltuaisten lukumäärän eron vuoksi ensin 4 eläintä jaetaan (lyhytaikainen vaihe 12 blastomeeria) ja sitten 4 vegetatiivista blastomeeria (vaihe 16 blastomerejä).
Seuraavat kaksi samanaikaisesti esiintyvää vakoa ovat leveyttä. Ylemmän leveysuran nopeampi läpikulku aiheuttaa 6 lyhytaikaista vaihetta, joka koostuu 24 räjähdyksestä. Alemman leveysuran läpikulun loppuun saattaminen johtaa 32 blastomeerin muodostukseen. Vaiheen 64 blastomeerien jälkeen, huolimatta siitä, että vakojen sekvenssi säilyy, rivin (2 - 4 - 8 - 12 - 16 - 24 - 32 - 64) geometrinen sekvenssi rikotaan. Murskauksen meridi- naalisten ja leveiden urien kulun rinnalla muodostuu myös tangentiaalinen ura, jonka seurauksena alkiosta tulee monikerroksinen.
Sammakkoeläinten alkioita, jotka sisältävät 16 - 64 solua, kutsutaan yleensä morulaksi, koska etäinen ulkoinen muistutus on Mulperinmarjaa (lat. Morum). 128-soluisessa vaiheessa ilmestyy selvästi erotettava blastocele ja on yleisesti hyväksyttyä, että alkio saavuttaa tällä hetkellä blastula-vaiheen, vaikka blastocoelin muodostuminen voidaan jäljittää aivan ensimmäisestä pirstoutumisjaosta (Golichenkov V.A.)
Kalojen murskaus
Polilekyyttiset munat
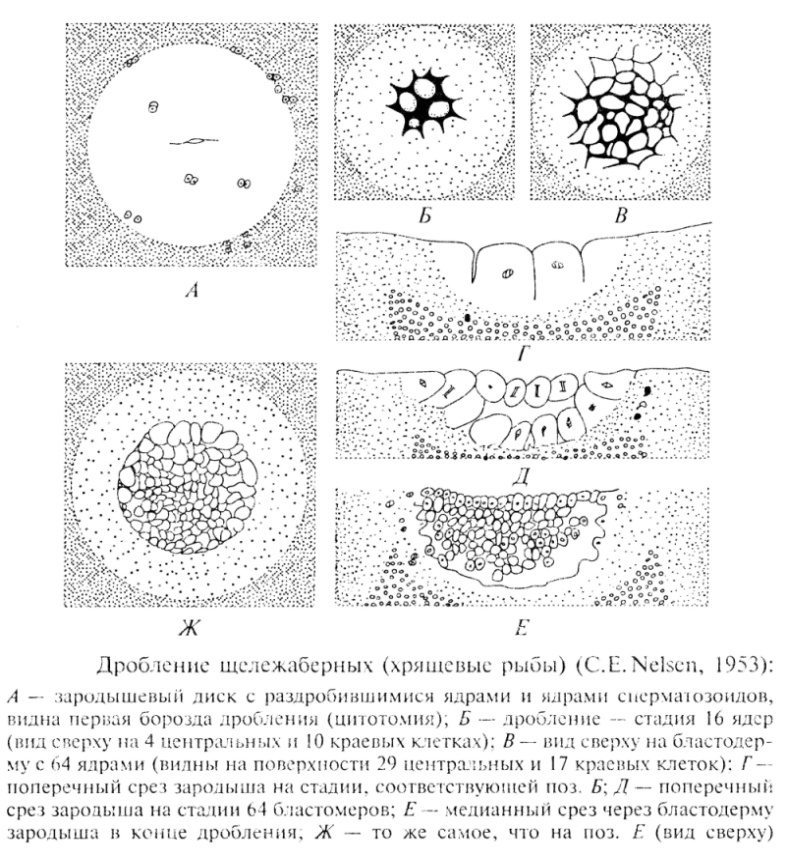
telolecithal. Juhlitaan
täen polyspermiaa. Usko siihen
siittiöt, jotka eivät ole fuusioituneet
naisen pronucleus, pitkä
varastoitu keltuainen kerros
munat, jotka rajoittuvat blastodermiin,
ja jopa osallistua kierrätykseen
keltuainen. Diskoidinen murskaus.
Murskauksen viides jako
erottaa joukon blastomeereja
keltuainen. Reunapuhaltimet
ja blastodermin juuressa olevat blastomeerit pitävät kosketuksen keltuaiseen. Kuusi murskausjakoa on enemmän tai vähemmän tahdistettu, minkä jälkeen synkronointi katkeaa. Seurauksena on, että blastodermin pohjaan muodostuu periblasti tai merosyyttikerros. Murskauksen seurauksena muodostuu diskoblastula, jota ulkoisesti rajoittaa tiiviisti kytkettyjen yhtenäisten solujen kerros. Ajan myötä blastulan rakoinen onkalo kasvaa, ja suuressa määrin blastodiscin yhdessä reunassa. Uskotaan, että erot blastocoelin koosta (tai toisin sanoen solujen pakkaustiheys blastodermin eri osissa) määräävät tulevan alkion akselin ja kahdenvälisen symmetrian - laajennetun blastocoelin alue vastaa alkion kaudaalista päätä.
Murskaus matelijoissa
Muna murskataan matelijoissa epätäydellisesti, diskoidisesti. Koska munan hedelmöitys tapahtuu munanjohdon ylemmässä kolmanneksessa, murskaaminen alkaa munasolun liikkuessa munasarjaa pitkin ja kun munia munitaan, alkio on joko diskoblastulan tai varhaisen gastrulan vaiheessa. Monissa matelijoissa, kun muna on pitkittynyt muna-olosuhteissa, muodostuu uusi organismi, joka syntyy tai poistuu munasta heti muninnan jälkeen (eläinperäinen lisko, virukset).
Diskoidinen murskaus johtaa diskoblastulan muodostumiseen. Blastulan katto koostuu pienistä blastodermisoluista, joita kokonaisuutena kutsutaan blastodisc-soluiksi, ja blastulan pohjan muodostaa katkeamaton keltuainen massa. Osa alkion levyn alla olevasta keltuaisesta resorboidaan ja sub-alkion onkalo muodostetaan raon muodossa blastodisc: n ulko- ja sisäkerroksen väliin.
Lintujen murskaus
Lintujen murskaaminen ja muodostuminen tapahtuu, kun muna liikkuu munasarjoa pitkin, ts. samanaikaisesti ympäristöönsä tertiäärisillä kuorilla. Murskausaika kananalkussa on 22 tuntia.
Tsygoottien murskaaminen lintuissa on epätäydellistä, diskoidista. Vain eläinnavalla sijaitseva alkiolevy murskataan, sillä sen pinta ja tilavuus on merkityksetön verrattuna keltuaisen murskaamattomaan massaan. Kolme ensimmäistä murskausuraa ovat säteittäisiä, jotka vastaavat lansetin ja sammakkoeläinten meridionaalisia uria. Sitten ilmestyvät leveys- ja tangentiaaliset urat (kuva 2).

Kuva 2. Kanan diskoidinen murskaus (Dondua, 2005):
A - 2 blastomeeria; B - 4 blastomeeria; B - 8 blastomeeria; G - 16 blastomeeria
Diskoidisen murskaamisen aikana, blastodisc-reunalla ja sen alla jäävät kaikki keltaiset kelluksesta epätäydellisesti erotetut aika-solut (periblast), josta keskimmäiset räjähdysprosessit on erotettu. Murskaus lintuissa on terävästi epätasaista ja vääriä. Alkaen kolmannen ja neljännen uran läpikulusta muodostuu erikokoisia soluja, joilla ei ole sääntöjenmukaisuutta ja pysyvyyttä sijainnissaan.
Muninta tapahtuu muninnan aikaan noin 14 tsygootin jakautumista, minkä seurauksena äskettäin munitun munan blastoderma sisältää noin 60 tuhatta solua. Seurauksena on, että muodostuu monikerroksinen blastodisc, joka koostuu epäsäännöllisen muotoisista soluista, jotka ovat lähekkäin toisiaan ja sijaitsevat katkeamattomalla keltuaisella (kuva 3).

Kuva 3. Kanan sukusolujen muodostuminen (Dondua, 2005):
A - murskauksen myöhäinen vaihe; B - ennen mahalaukun aloittamista:
1 - blastomeerit; 2 - epiblasti; 3 - synnytyksen ontelo; 4 - keltuainen
Blastodisc-solujen ja merosyyttien vaikutuksesta, osa keltuaisesta nesteytyy keltuaiseen, muodostuu pieni tila,
täynnä nestettä - alkion ontelo. Tässä vaiheessa kaksi blastodisc-aluetta on selvästi erotettavissa: kirkas kenttä keskellä (pellucida-alue) ja tumma kenttä reunalla (opaca-alue). ( Golichenkov, V.A.Työpaja)
Kirkkaan kentän ulkonäkö johtuu siitä, että blastodiscin keskellä
alkio käyttää ensinnäkin alkioita, minkä seurauksena muodostuu sub-alkion rako ja tämä koko alue näyttää läpinäkyvältä. Blastodisc-reunan reuna-alueen solut menevät päällekkäin keltuaisen kanssa, joten alkion tämä osa näyttää tummalta. Golichenko, V. A)
murskaava umlekopitayuschih.
Pian sen jälkeen, kun tsygootti on muodostunut, alkaa sarja mitoottisia jakautumisia, joita genomi valvoo tiukasti ja joita kutsutaan fragmentoitumiseksi; tämä prosessi alkaa useita tunteja hedelmöityksen jälkeen, jopa munasoluissa. Jokainen jako kestää 12 - 24 tuntia. Tsygootti on jaettu blastomeerisoluihin tietyssä sekvenssissä. Ensimmäinen jakautuminen tapahtuu tasossa, joka kulkee munan molemmat navat, joten pallomainen sigotti on jaettu kahteen pallonpuoliseen blastomeeriin. Jaon syntyvä halkeama ei sijaitse satunnaisesti, vaan riippuen siittiöiden tunkeutumispaikasta ja myöhemmästä sytoplasman uudelleen jakautumisesta. Murskauksen toinen ura kulkee kohtisuorassa edelliseen nähden. Saadaan neljä blastomeeria, ryhmiteltynä radiaalisen symmetrian periaatteen mukaisesti, sitten ne jaetaan jälleen puoliksi muodostamalla kahdeksan blastomeeria. Jatkuvaan blastomeerien fissioon on ominaista fissio- tasojen vuorottelu - toinen blastomeereistä jakaa päiväntasaajan tasossa, toinen meridionaalitasossa. Tätä nisäkkäille tyypillistä jakautumista kutsutaan vuorottelevaksi. Lisäksi blastomeerit eivät jakaudu samanaikaisesti, ja tämän seurauksena solujen lukumäärä ei selvästi kasva 2: sta 4: een ja sitten 8: een. Tietyissä ajankohtina alkioissa on pariton määrä blastomeerejä.
Kukin murskauksen aikana muodostuneista soluista on noin 2 kertaa pienempi kuin äiti. Murskausjakojen välisellä ajanjaksolla vaiheiden välistä solukasvua ei tapahdu, ja siksi kaikkien solujen kokonaismassa pysyy suunnilleen yhtä suurena kuin munan massa.
Kuva 1. Nisäkkäiden alkion kehityksen varhaiset vaiheet:
I - murskauksen alku;
A - hedelmöitys; B, C, D - blastomeerit;
II - morulan muodostuminen - tummien solujen asteittainen likaantuminen valolla;
III - blastosystojen muodostuminen:
A on onkalon muodostumisen alku;
B - sukusolmukkeen erottaminen trofoblastista;
C - sukusolmun muuttuminen sukusuojaksi (diskoblastula);
1 - trofoblastit; 2 - embryoblast.
Jakamisen seurauksena muodostuu monisoluinen alkio, joka muistuttaa ulkonäöltään vadelmamarjaa ja jota kutsutaan morulaksi. 8-soluisen morulan vaiheessa, joka on ominaista vain nisäkkäille, tapahtuu 6 elastomeerin merkittävä konvergenssi. Tässä tapauksessa solujen välillä syntyy tiukkoja kontakteja, jotka sallivat pienten molekyylien ja ionien siirtymisen solusta soluun. Tätä ilmiötä kutsutaan tiivistykseksi ja se myötävaikuttaa alkion erilaistumiseen ja ravitsemustoiminnon omaavan trofoblastin sekä sisäisen solumassan, joka synnyttää itse alkion, erottumiseen. Tiivistysprosessissa alkion solujen plasmakalvon yksittäiset osat alkavat liikkua eri, tiukasti geneettisesti määritettyihin suuntiin. Solumembraanien koostumukseen osallistuvat proteiinit osallistuvat näihin prosesseihin, ja kun esiintyy blastomeerejä toisiinsa kiinnittäviä mikrovilejä, niiden sytoskeleton muutokset.
32-soluisessa morula-vaiheessa koiran alkio saapuu kohtuun. Tämä tapahtuu 7. - 8. päivänä, mikä on paljon myöhemmin kuin muilla nisäkkäillä. Tänä aikana kehitys pysähtyy ja alkioita kuolee jakautumishäiriöillä, jotka johtuvat sekä perinnöllisistä että ympäristötekijöistä. Siten perinnölliset ja ympäristötekijät ovat aktiivisia luonnollisen valinnan tekijöitä.
Jakautumisprosessin aikana saadut räjähdyssolut sijaitsevat reuna-alueilla, ja vähitellen morulan tiheän solumassan keskelle muodostuu onkalo (räjähdysväli) ja solujen itämäinen kerääntyminen - embryoblast, jota muuten kutsutaan itusolmuksi, ja syöttösolujen ympäröivä kerros - trophoblast erotetaan. Alkion runko muodostetaan myöhemmin alkion räjähdyksestä. Trofoblasti toimii ravitsevana lehtenä varhaisalkioille. Tätä alkion kehitysvaihetta kutsutaan blastosysteeksi.
Tässä vaiheessa oleva blastosysta koostuu trofoblastisolujen ulkokerroksesta ja sisäisestä solumassasta, pallomaisesta soluklusterista, joka on kiinnitetty sisäpuolelta yhteen trofoblastin napoihin ja edustaa materiaalia alkion rakentamiseksi. Solun tuleva sijainti sijainnissa alkiossa tai trofoblastissa määritetään tiivistysprosessissa, kun solut esiintyvät joko alkion pinnalla tai sisällä.
johtopäätös
Viitteet
Almazov I. V. Atlas histologiasta ja embryologiasta./ Almazov I. V., Sutulov L. S M .: Medicine, 1978.
Belousov L.V. Yleisen embryologian perusteet. - Moskova: Moscow University Press: Science, 2005.
Golichenko, V. A. Embryologian työpaja / V. A. Golichenko,
Gilbert S. Kehitysbiologia. T. 1. M .: Mir, 1993.
Goličenko, V. A. Embryologia / V. A. Goličenko, E. A. Ivanov,
Dondua A.K. Kehitysbiologia. Vertailevan embryologian alku /
Carlson B. M. Patten-embryologian perusteet. T. 1. M .: Mir, 1983
Knorre A. G. Lyhyt kuvaus ihmisen sikiöstä. M .: Medicine, 1967
Tokin B.P. Yleinen alkiotutkimus: Oppikirja. biol. spec. un-ta. - 4. painos, tarkistettu. ja lisää. - M .: Korkeampi. school., 1987. - 480 s.
Sotskaya M.N. Koirien sukutaulujen jalostus. / Sotskaya M.N., Moskovkina N.N. - M .: Aquarium-Print LLC, 2006. - S. 95-109.
Sze, S. M., Puolijohdelaitteiden fysiikka, John Wiley ja Sons, N.Y., 1981.